
by Raviv Ganchrow and Carsten Stabenow
Whale Refractions engages with circuited operations (Ganchrow 2024) and diffuse continuities discernible in mammalian thalassic agency. It addresses complex planetary dynamics, involving coupled organic-inorganic oscillations cutting across temporal, geological and technological domains. Spatial-material dimensions of those operations come into sharpened focus while refracting through Magallanes contexts and sub-Antarctic tectonics.
Whale Refractions adopts sound as a guide on a journey through planetary ‘heritage at large’, tracing shifting relations in human-whale attentions, late Gondwana geology, cetacean and auditory morphogenesis, Eocene global warming, oxide-based recording techniques and piezoelectric oceanic surveillance, all entwining through a set of undulations typical to the Age of Mammals.
Ultra-broadband Listening
Following-up on experimental underwater spatial monitoring in Magallanes, during the austral summer of 2025; testing of an upgraded system is currently underway, the following sections discuss preliminary insights and discoveries from the recent work. The newly designed system provides an extended monitoring range — from below 1 Hz till above the upper limits of human hearing*. It incorporates experimental, purpose-built hydrophones (underwater microphone), developed for Whale Refractions research. This ultra-broadband transduction tunes into the full gamut of sub-Antarctic marine activities. From ultraslow seismic modulations, through midrange marine mammal and fish vocalizations, up to the high-pitched clicking of crustaceans and glacial melt. The system offers a continuous polyvalent audio feed, open to experimental approaches to listening and open-ended analysis.






Songs in Magallanes
A whale ‘song’ is a technical term applied to distinctive patterns of sounds with repeating elements and hierarchical structure produced by a lone vocalizing member of a pod (Schulze et al. 2022; Payne and McVay 1971). There remains lively scientific debate regarding the meaning and function of whale songs —abundant in warm-water equatorial breeding grounds— together with a renewed interest in the specific historicity and techniques entangled with hearing whale vocalizations as songs. That list includes, but is not limited to, 1970s environmentalism, Cold War submarine surveillance and interspecies cultural imaginations (Burnett 2012; Bakker 2024; Steingo 2024).
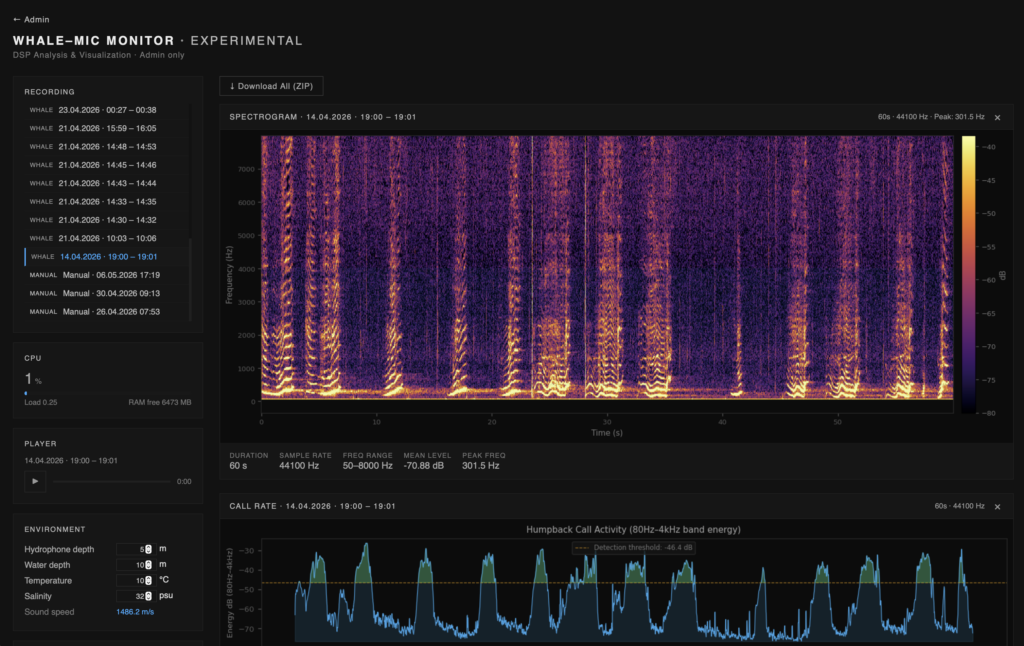
In the past decade, humpback whale songs have also been observed in southern hemisphere feeding grounds, where they tend to appear towards the austral fall. After prolonged feeding activities, fall is the season in which the pod prepares its annual migratory return (Schall, et al. 2021; Español‐Jiménez and Van Der Schaar 2018; Stimpert et al. 2012). Up until now, there have been no documented songs in the Straits of Magellan. On April 14th, 2026, shortly before 4 AM local time, a whale song was picked up near Isla Rupert. The signal was broadcast over our recently installed experimental hydrophone in the bay of Isla Carlos III, relayed via satellite and recorded via the online live stream in Berlin and Amsterdam. The audio recording vividly rendered a vocalization sequence consisting of humpback whale screeches, moans, purrs, grunts, groans and trills. The roughly 6-minute song is the first documented recording of its kind in the Magallanes region.
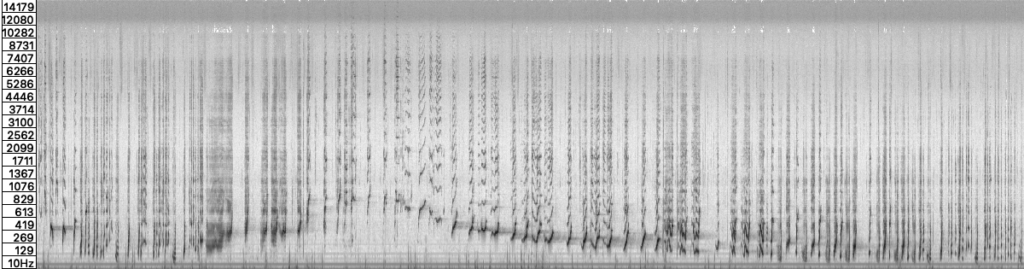

Preliminary analysis suggests a multi-themed humpback song with distinctive units in nested patterns. The recording consists of 120 vocalization units (discrete acoustic events), structured in phrases and themes. Unit lengths range from 9.5 seconds (first third) to 0.3 seconds (final section). Inter-unit gaps range from 4.5 seconds to 0.5 seconds. Though the term — song — is somewhat loaded with normative anthropomorphic musical meanings that require some unpacking. Especially considering the pitch analysis of a song is biased towards its self-contained note-to-note melodic relations, diminishing a song’s linguistic heritage not to mention its environmental agency. Historical analysis of humpback songs reads gaps between units as silent, excluding the emplaced and spatial dimensions of the singing. Environmentally active components of vocalization are precisely the aspects to which Whale Refractions research is currently attending. For example, linguistic dimensions of humpback songs, unlike birdsongs, have marked similarities with human speech syntax, blurring distinctions between speaking and singing (Arnon et al. 2025; Suzuki et al. 2006). What if such definitions were expanded to include spatially situated conditions of vocalization? Maybe the whale song considered as a kind of speak-sing-place? The notion of sung emplacement also reframes an interspecies component of whale vocalizations that under certain conditions was astonishingly audible above water from our basecamp on Carlos III, suggesting there could be reciprocal vectors of musicing whales in Fuegian cultures that confound western classical notions of representation.
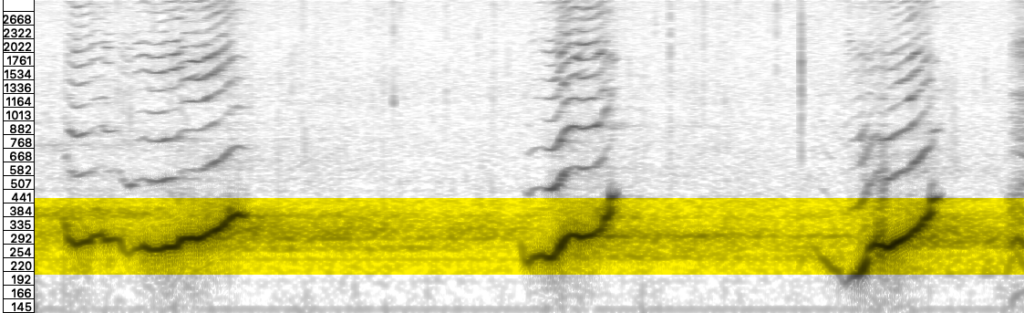
In the recording, song units consist of grunts, honks, and modulated tones with an energy envelope of increasing intensity that ceases abruptly. Units often begin with a noisy-spectra gurgle honing in on distinctive frequencies whose audible hues extend and transform in the timbral shimmer of environmentally active reverberations. It is as though the unit energy profile (amplitude envelope) is structured for producing reverberant traces. With that framework in mind, the inter-unit gaps are far from silent, but rather teeming with activity, potentially adapted to echoing returns. Inter-unit gaps provide intervals where the environment rushes in, conveying crucial source-environment interactions. One notable sequence in the song consists of upwards sliding pulse trains, incrementally downward stepping in tone. Not unlike the ‘megaclicking’ observed in north Atlantic humpbacks, though more rich in timbre (Stimpert et al. 2007). The down-pitching stabilizes on a spectral center around 270 Hz in the second half of the song, a spectral zone that is exceptionally spatially responsive to the calls in the waters off the coast of Isla Rupert. Stated succinctly, the whale song frequencies and wavelengths activate resonance structures of the near archipelago.
Songs are relatively rare occurrences in a broad range of non-singing social sounds humpbacks produce such as gurgles, groans, snorts and thwaps observed throughout feeding behaviors. Foraging sounds occurred in groups of 2-6 humpbacks and were more prevalent when the groups were larger. Socializing sounds were observed across members of the pod, including young calves whose vocalizations are substantially higher pitched. Behaviors such as synchronic diving and dive-duration coordination may also have sonic cues that require cross-referencing of video footage with the hydrophone recordings.






Archipelago Reverb
Audio analysis reveals extended reverberations in the archipelago, possibly as long as 15 seconds, which can be attributed to the surface hardness and cavernous properties of post-glacial submerged basins, characteristic of the channels surrounding Isla Carlos III. The archipelago functions like a gigantic set of interlinked chambers. But also, interestingly, chambers whose reverberation times ring differently in accordance with bandwidth, revealing prominent resonance signatures of the underwater morphology that, incidentally, are particularly responsive to the humpback whale calls. For example, the sweeping glissando section in the recorded whale song eventually settles on a reverberant zone of ocean spectrum, spanning a range from 130-400 Hz. The reactive zone is then repeatedly revisited in units from the second half of the song, centered around 270 Hz – precisely the median of the reactive bandwidth.
The bandwidth from 1000-3000 Hz is notably saturated in nearly all the recordings. This could be attributed to wave-bathymetry structural interactions and requires further analysis. To a lesser extent, the bandwidth from 4500-8000 Hz displays intermittent saturation likely due to ocean surface interface interactions and wind and precipitation. Conversely, the 200-1000 Hz spectral band shows relative quiet. It is also highly reactive to the fundamental frequencies and lower partial of humpback vocalizations, possibly a significant bandwidth in their auditory system.
Detailed whale song recording analysis reveals a reverberation for the eastern bay of Carlos III estimated at 8-15 seconds (bandwidth-dependent RT60). Deep Chilean channels are largely unmeasured, but compare those estimates with Norwegian fjords (4-12 second reverberation), or with the benchmark reverberation of modern concert halls of around 2.3 seconds. Every channel or fjord in the sub-Antarctic acts as a waveguide, with its own unique sound-refractive characteristics derived from the channel’s cross-section geometry, the salinity and temperature of the water, the sum of which are also influenced by ongoing glacial and tidal flows. In fact, the entire Straits of Magellan likely has its own signature resonances and a cutoff frequency that could be acoustically modeled with accurate bathymetry.
Song Echo Ranging
Echolocation in baleen whales remains controversial amongst marine zoologists. Although some researchers have attributed echolocative functions to the whale song (Mercado 2018), at this time there are no known biological mechanisms to back up such claims. Nevertheless the consistent gaps between sections of the vocalization indicate the potential humpback monitoring to the echo returns in the fjords. Mapping the predominant reflections, derived from intervals of the recorded song, provides a time-map of near bathymetric features. Interestingly, the inter-call timing (median 2.8 seconds) correlates with a round-trip distance of just over 4 km, which falls within the range of most of the near islands and bathymetric features in the vicinity of the singing whale (approximately 2 km distance between Isla Carlos II & Isla Rupert). This is consistent with the hypothesis that the whale’s inter-call spacing is potentially adapted to the channel geometry, where humpbacks are listening to qualities of their echo-returns. Our recording provides compelling evidence for spatially responsive features of humpback singing that requires more careful examination.
Preliminary analysis of the reverberation structure, over several consecutive calls, reveals a pattern of approximately two dozen prominent peaks, corresponding with echo-returns from bathymetric features (such as near islands, steep underwater slopes, ridges, knolls, and basin walls) ranging between 39 meters and 2044 meters, some with repeating echoes in harmonic series. A comparison was made with a section of the same recording that contained no vocalizations to determine the noise floor in the bay. At lower frequencies (below 200 Hz), ambient noise masks accurate estimates of the reverberation, however the ‘silent’ signal also retrieved 14 of the 20 reflections appearing in the vocalized section of the recording – confirming predominant structures of the bay.
Cross-referencing the echoes with a bathymetric map reveals the spatial features. The most distant prominent reflector, calculated at a distance of 2044 m from the hydrophone, most likely corresponding to a steep concave incline of Rupert Island (reflector T). Reflectors P and Q, only activated during vocalizations, indicate the whale position in the waters between Carlos III and Rupert islands, slightly to the north of the hydrophone position. Most reflective sources in the near-field (0-900m) remain spatially unresolvable from a single recording. One exception is a reflector estimated at 134m, corresponding with two plausible boundary conditions of the shallow bay of the hydrophone. Difficulty in determining near-field reflections may be due to the bathymetric quality of the commercially available map applied in this preliminary survey, especially considering the spatially unresolved zone near Carlos III. The mid-basin region and Rupert’s landmass has better detail, providing at least six spatially resolvable echo ranged candidates, corresponding with ridges, knolls and concavities of underwater topography.

Vortex Currents
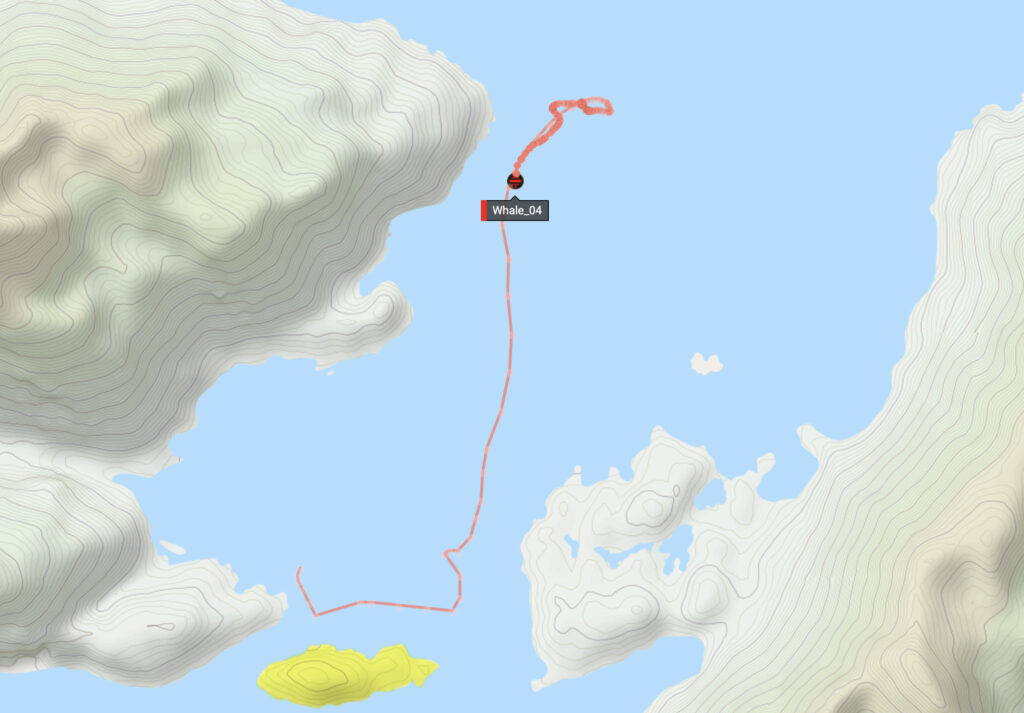
In parallel with the fixed monitoring, testing continued with the experimental whale-binaural buoys, the Ocean Current Tethered Hydrophone Array (OCTHA). Whale 2 & 4 OCTHA sonobuoys were deployed simultaneously, towards the southern end of the Barbra Channel, south of the Straits of Magellan, though Whale_2’s GPS signal jammed rendering Whale_4 the only mapped trace. Eight hours later, both OCTHA sonobuoys were retrieved, still in close proximity to one another, apparently after having followed nearly an identical course of movement. The vortices and labyrinthine waterways of the channel, during tidal transition, produced a striking cyclic drift pattern roughly tracing a figure-8.
The abundance of humpbacks in Barbra Channel (increasingly frequented by Punta Arenas whale watching industries) is potentially linked the specific properties of the channel. The unusual current patterns with strong vortices likely contribute to nutrient upwelling and pockets of rich feeding. With motors off, our 45 meter-long iron-hull boat Estoa was thrust in the currents like a spinning top. Underwater echoes in this section of the channel are exceptionally pronounced, suggesting an equally complex, cavernous deep-water topography. Preliminary sonar mapping of the island obstructing the south end of the channel, where vortices prevail, indicate a complex, rocky rippled basin structure with rapid drop-off to depths exceeding 100 meters. Further bathymetry analysis is required to map interlinks between ocean current movements, nutrient upwell and archipelago reverb.








Whale Representations
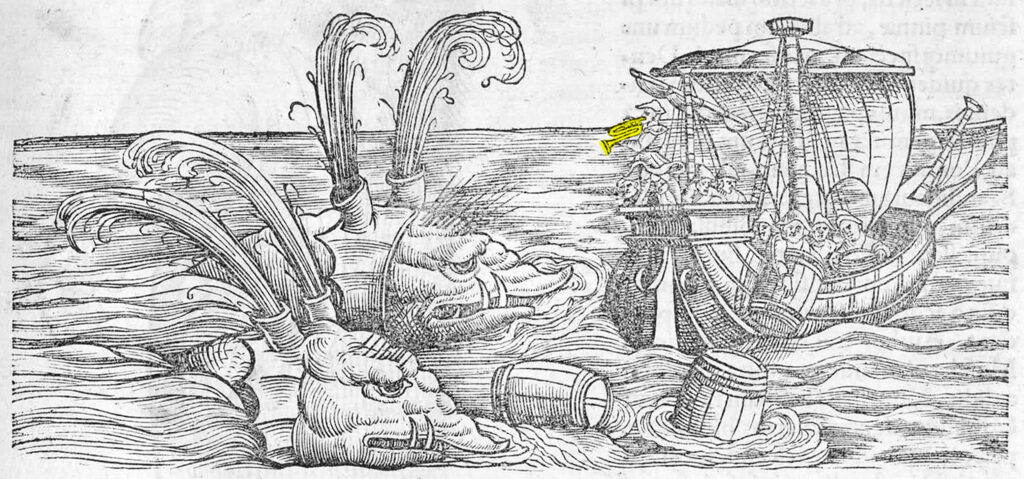
Early accounts of passage through the Straits of Magellan noted the abundance of cetaceans. Whales can be found throughout mythologies of Chilean coastal communities. Whales are integral to Fuegian cultures, reflected in Yahgan, Selk’nam and Kawésqar folklore and music. Whales are evoked in Selk’nam song and Hain ceremony body paint patterns, though they rarely appear as representational depictions, with the notable exception of the El Médano rock paintings of coastal Atacama (Niemeyer 2010). Conversely, whale representations have always appeared in the fringes of colonial and naval maritime maps. Though often depicted as monstrous apparitions capable of swallowing boats. Edges of maps chart the limits of knowledge, spilling over into shared speculative imaginations that are also informed by whale vocalization heard by mariners transducing through hulls of wooden ships, attributing them to mythical underwater creatures.*** Herman Melville traces a partial genealogy of those cross-cultural imaginaries in Moby-Dick’s chapter titled ‘Of Monstrous Pictures of Whales’. Four depictions of such aquatic behemoths appear alongside a mermaid and flock of flying fish in an early map of Patagonia, that includes Tierra del Fuego. The map by Alonso de Ovalle, a Chilean Jesuit priest, was incorporated in a book published in Rome in 1646, aimed at recruiting new missionaries for the church’s southernmost outposts.

Although there are no records of whales downing ships in Tierra del Fuego, a humpback did gulp down and promptly spit out a packrafter last year in the straits, at the bay of eagles (Bahía El Águila). The event was captured on action-cam, its refractions via social media and news outlets, circumnavigated the globe. During a Terra Ignota survey of Bahía El Águila, our camera, too, inadvertently captured the two packrafts in the bay. Moments before the yellow vessel was briefly consumed.
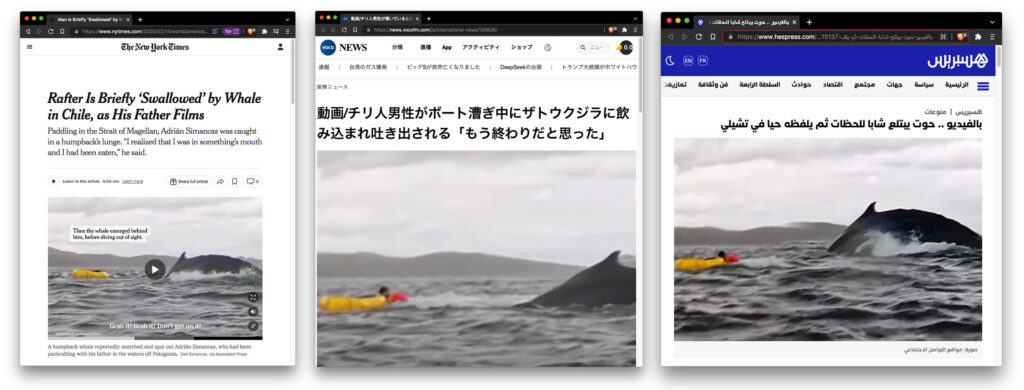
Incidentally, the miraculous event occurred directly above a mass grave of whales. In the early 20th century, Bahía El Águila housed a whale oil processing facility run by the Sociedad Ballenera de Magallanes (the Magellan Whaling Company), the first modern whaling facility in Chile founded on a skillset, technology and a whaling ship (Almirante Montt) imported from the Norwegian town of Sandefjord. El Águila’s shallow bay is still strewn with countless whalebones. Possibly the foraging humpbacks took notice? During our brief visit to the cove, with the guidance of Miguel Cáceres of Museo De Historia Natural Río Seco, we recovered a dozen eroded whale ears (tympanic bullas and petrosal/periotic bones) washed up along coast. This summer, while deploying hydrophones near Carlos III, we were warned by locals that our yellow packraft is particularly appetizing for humpback whales.




Distant refractions from former missionary efforts were also present in the Chilean packrafter, Adrián Simancas’ account. In his interview on CNN (14-02-2025), Simancas likened the experience to that of Pinocchio & Geppetto trapped in the belly of a sea-monster. A plot Carlo Collodi’s book ‘Le avventure di Pinocchio’ (1883) refracted from an earlier book — the bible — recounting experiences of the prophet Jonah (c. 2800 BP) who spent three days and three nights in the belly of a large great sea creature (דג גדול, dāḡ gāḏol), literally translated as ‘large fish’, before escaping by divine intervention. The migration of narratives and symbols, over geographies and across time, attest itineraries of social transduction. The tale of ‘Jonah and the Whale’ is shared by Judaism (יוֹנָה Yōnā), Christianity (Jonas) and Islam (يُونُس بْن مَتّى، Yūnus ibn Mattā), though it’s only recently that impression of the colossus fish have been grafted onto whales. Today Jonah’s whale most often appears as a sperm whale – a deep-diving cetacean that feeds on giant squid and possibly the only species capable of swallowing a human. Disney chose a sperm whale for the rendition of Monstro in their version of Pinocchio (1940). Other baleen whales would barely manage swallowing an arm. A humpback’s esophagus, for instance, is constrained to the size of a fist. And anyhow, its only humans that have ever eaten whales, never the other way around.

Beside the monstrous whales on de Ovalle’s map are other questionable imaginaries, such as depictions of Tierra del Fuego Island inhabitants as bow-wielding tailed ‘men of Palmaris’ of the ‘straw and mud clothed’ inhabitants of Cape Horn – depictions of Selk’nam and Yahgan territories respectively. The map also bears the Dutch name of Cape Horn: ‘Hoorn’. Capo Hoorn, named after the Dutch city of Hoorn, was the southernmost headland of the Tierra del Fuego archipelago. At the time, part of a new maritime rout charted three decades earlier by mariners from Hoorn, seeking to evade Dutch East India Company (VOC) taxation controlling the straits. Up until then, the Straits of Magellan presented the only navigable Pacific-Atlantic crossing, the discovery of the southern circuit significantly rerouted maritime traffic in the region. The digging of the Panama Canal (operational since 1914) provided a shortcut, remapping the centuries old crossings via the straits, Drake Passage and Beagle Channel.
Close by the controversial depictions of Yahgan and Selk’nam inhabitants in the de Ovalle map, two proto-whales also appear, one of which directly links up with the story of Jonah, this time via the Flemish engraver Anton Wierix II whose lithograph of the biblical fable, completed 61 years earlier, apparently served the basis of de Ovalle’s Fuegian variant.
At first glance the Fuegian whales match the fanciful sea monster trope, composed of a hodgepodge of animal parts with gigantic fish-scaled flukes. Though upon closer inspection they disclose the two primary variants of whale types described as far back as Aristotle’s ‘Historia animalium’ (History of Animals c. 2,350 BP), discerned in mouth structures that differentiate the baleen whales from their toothed cousins. The Mysticete (baleen bearing) and Odontoceti (toothed) evolutionary split is estimated to have occurred around 40 million years ago, 10 million years after initial immersion. Conventional distinctions between echo-locating toothed cetaceans and filter-feeding baleen whales also defines the two remaining branches of whale evolution. However, the spectrally calibrated and spatially-response humpback calls observed in Magallanes troubles that binary that relies on the hackneyed metaphor of clearly discernible branches in Darwin’s tree of life. Perhaps echoes form beneath Darwin’s boat, in the nearby Beagle Channel, can help remedy the metaphor — displacing the object bias of ontology with a vibrant ontogenesis of becoming.
Multiple species from both baleen and toothed variants can be found throughout the sub-Antarctic archipelago, though often constricted to specific regions, features that apparently did not pass unnoticed. In Magellan’s original account of navigating the straits, the ship anchored at a place they named ‘River of Sardines’ – a staple humpback food.

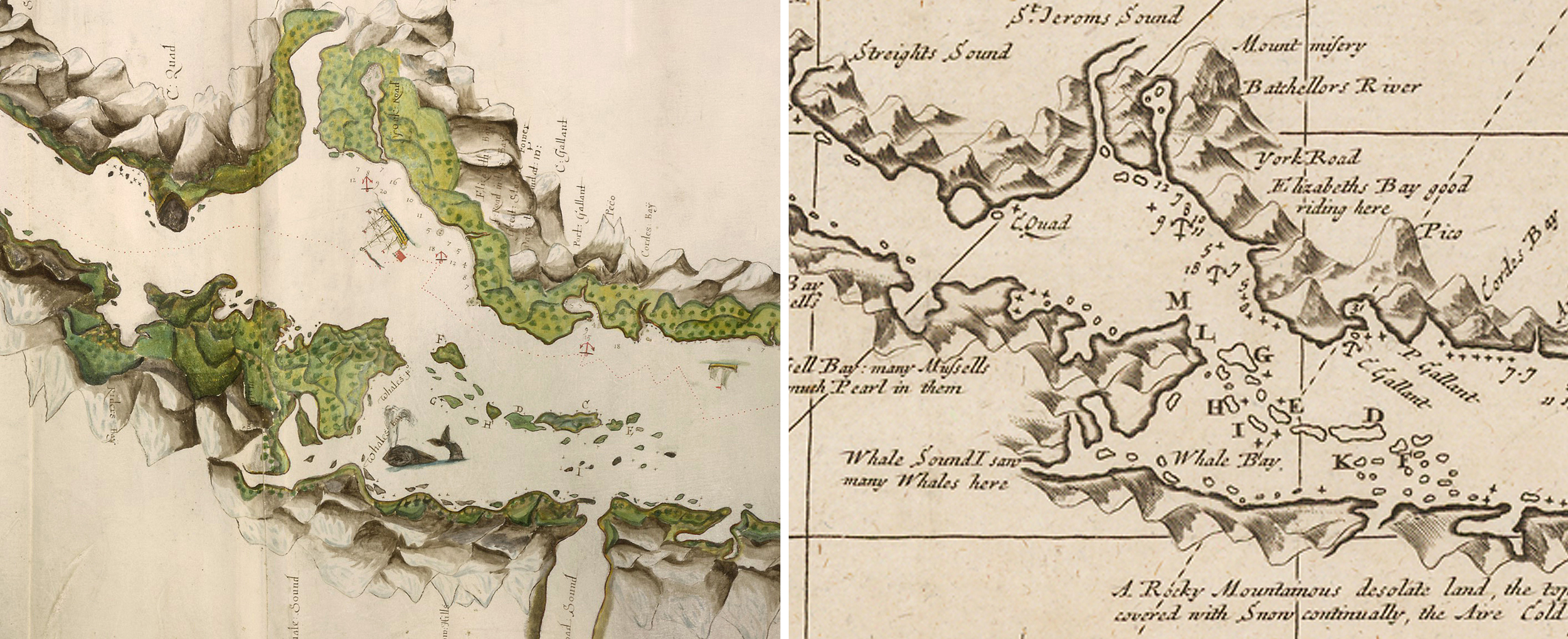
In stark contrast, John Narborough’s painted map of the Straits of Magellan from 1670 has a rather convincing depiction of a baleen whale, with the exceptions of its double-spout blowhole, similar to de Ovalle’s rendering, likely tracing back to Conrad Gessner’s double piped ‘Monstrous Whales’ that echoed 16th century sentiments of aquatic giants that were simultaneously demonic and divine.
Narborough’s whale is depicted swimming near a prominent peninsula in the Straits of Magellan (now confirmed to be Carlos III Island), at an area designated with the title ‘Whales Bay’. A printed version of the map from 1694 is more instructive, including the designation of ‘Whale Sound’ with its adjoining channel to Whale Bay as well as an additional inscription ‘I saw many Whales here’, with the exception of the whale drawings, absent in the revised map.
Monitoring Agency – Whale-mic Isla Carlos III
In the late 1990s and early 2000s an adventurous group of Magallanes marine zoologists, biologists and local mariners studied a prominent migratory destination of humpback whales in the southeastern Pacific, congregating annually in the fjords surrounding Isla Carlos III in the Straits of Magellan (Gibbons et al. 2003). The crucial discovery of the first known feeding ground for humpback whales along the Pacific coast of South America lead to the establishment of Chile’s first protected marine zone: the Francisco Coloane Marine Park in 2003. Since that time, the whales have been observed returning to these Austral feeding-grounds for over twenty years.
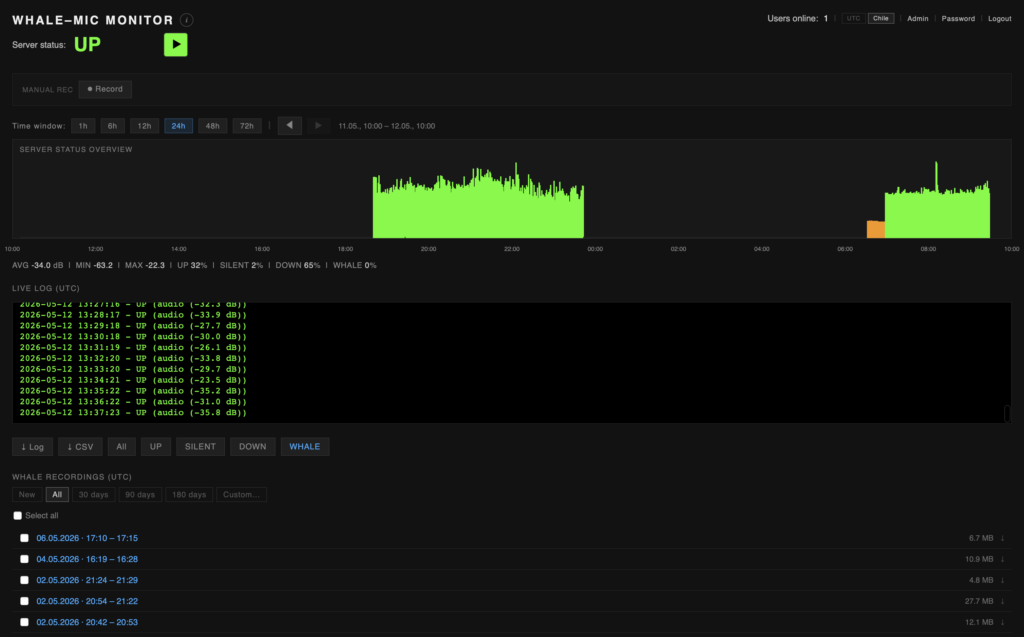
In January 2026, Raviv Ganchrow and Carsten Stabenow, with the assistance of Whale Sound – custodians of the marine station and founding members of the park — installed the first semi-permanent underwater microphone off the coast of Isla Carlos III. For the past two summers, Whale Sound has collaborated on the Whale Refractions project, commenced at the Centre for Advanced Study inherit at Humboldt-Universität zu Berlin, extended through ongoing work with the interdisciplinary research platform Terra Ignota. Transmission of the live-feed, realized in collaboration with Udo Noll and radio.earth, can be directly monitored here**:
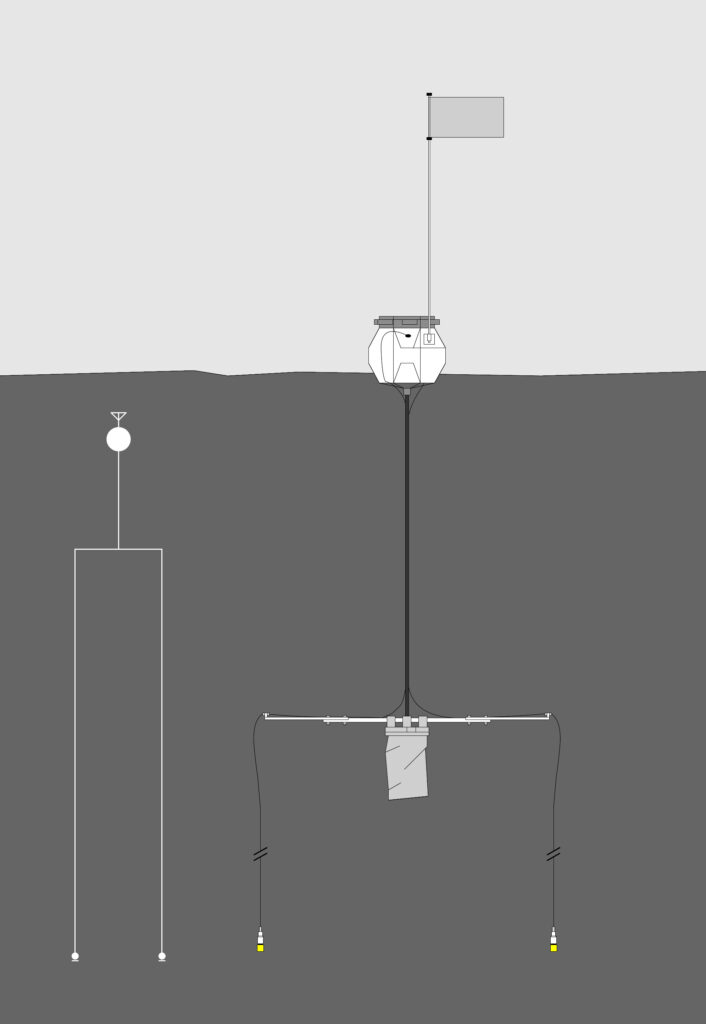
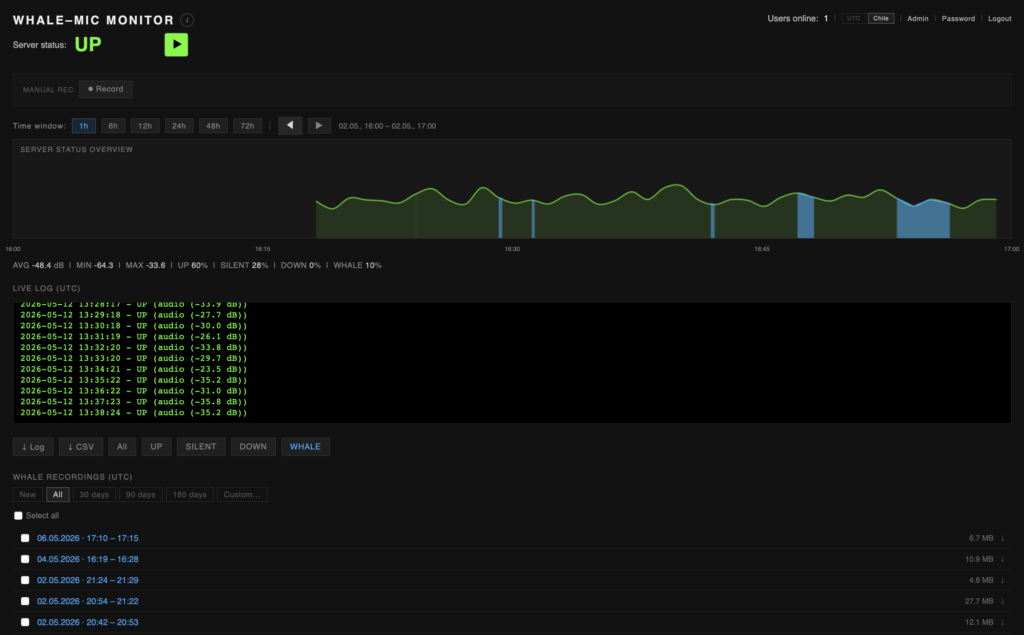
The hydrophone is moored in a shallow cove on the eastern coast Isla Carlos III, in the Fuegian fjords to the south of the Straits of Magellan (S 53°39.34122′, W 72°15.45183’). Near Whale Sound camp, 50 meters from the shoreline, at a depth of ca. 8 meter. The system consists of a phantom-powered custom-build hydrophone, with lead zirconate titanate piezoelectric element with an integrated high-Z amplifier circuit (developed in collaboration with Triton Audio, NL). The signal is relayed back to the camp over a submarine cable and routed online via satellite transmission. The entire system runs on solar power. The setup is a long-term prototype material test for a low-cost, ultra-broadband, semi-automated aquatic monitoring and recording system.






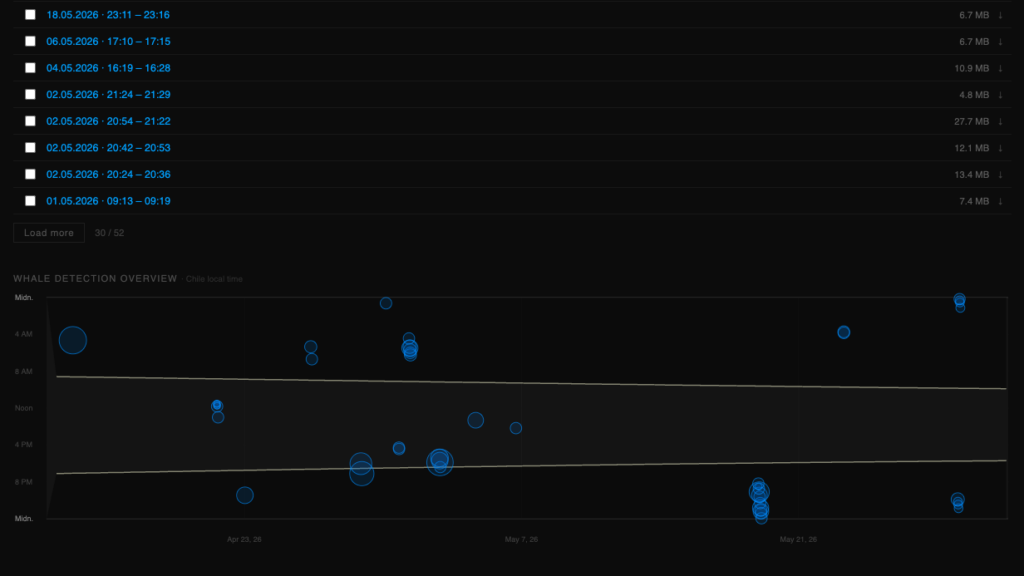
A dedicated automated monitoring platform is in development, analyzing the live signal for humpback vocalization detection, based on ML classifier via the TensorFlow Hub. Access to the platform and recording database can be granted to researchers upon request. (whalemonitor@terra-ignota.net)
Hearing Magallanes Contexts
Just how such open-ended listening redistributes the sensible — enabling immanent spaces of action — is yet to be determined. Though the monitoring inevitably participates in other shifting attentions to Magallanic sounds. For example, the troubling decline of Magellanic penguins on Magdalena Islands (Nahas Miranda 2026), with colonies extending into the Coloane park, whose diminishing squawks are potentially entangled with a tourist rush on Antarctica, shifting climates and emerging geopolitics of the thawing poles.
Similarly, the swarming sounds of Antarctic krill, Fuegian sprats and squat lobster are closely coupled to pressures from shifting climate and fishing industries that carry-over to the sounds of whales and other marine life dependent on them. Humpbacks apparently have a way of vocally signaling the changes, as anchovies transition to krill (Baisas 2025; Ryan et al. 2025). The Straits of Magellan hosts an immensely diverse, yet fragile biological web, that is also markedly susceptible to perturbations (Andrade et al. 2026). Climate variability affecting phytoplankton cycles, coupled with larger weather patterns such as El Niño carry over into the local ecologies (Hayward et al. 2025; Shabangu and Kowarski 2022; Schall et al. 2021).
Despite its difficult climatic conditions, the Straits of Magellan remains a strategic route for global navigation. A current concern is the rise in maritime traffic linked with booming Antarctic tourism, green-hydrogen production and increasing freight shipping. Accumulating engine noise in the exceedingly reverberant straits may be capable of hindering marine mammal (and other vertebrate/invertebrate) acoustic coordination and navigation. A recent survey of anthropogenic noise, effecting pilot whales in the Straits of Gibraltar, emphasizes the need for better acoustic understanding of sea mammal behaviors and general guidelines for managing increasing human activities in marine environments (Hegeman et al. 2026). Currently the Straits of Magellan is ranked as a global leader in fatal collisions with cetaceans (Toro et al. 2025; Guzman 2016), figures that are only set to rise with the introduction of new ports along the straits and recovering whale populations.


Eco-Links
What constitutes sub-Antarctic underwater ambiences and what does human aquatic-hearing entail? The live audio feed from Isla Carlos III hydrophone relays a flat-spectral-ontology, not favoring any species nor biased towards specific geological bandwidths. Signal relay, is a basic circuit function, to become accustomed to southern aquatic cycles. Broad-banded inclusivity encourages an open-ended listening, anticipating emergent modes of hearing and unforeseen spaces of action. Call it Eco-Links, homage to Marianne Amacher’s seminal City-Links #4 (Tone and Place, Work I), streaming live sounds from Boston harbor into Amacher’s studio/laboratory at MIT (1973-1976).
The Isla Carlos III hydrophone feed encourages recurrent (and prolonged) listening to the complex patterning of earth-bound interrelations. Subjecting attention to gradual tidal flows; sensing the shifts in precipitation; the patterned pattering of rainclouds, wind-swept waves and rhythmic clatter of seafloor gravel. Intimating the rubbing of kelp and crustacean clacking intermixed with the murmurs of fish and echoing reverb of marine mammal vocalizations; all occurring simultaneously and diffused within one another; signature Magallanes sounds that themselves have endured and transformed across eons of relational interactions.
Research conducted by Raviv Ganchrow and Carsten Stabenow
* Signal relay from hydrophone via satellite transmissions and across multiple servers to your computer browser inevitably reduces signal bandwidth to a range of approximately 5-11,000 Hz due to complexities of the transmission chain.
** Due to the limited energy resources on the island and increasingly darkness as winter approaches, the server is increasingly inactive as the season progresses.
*** The mapping section of Whale Refractions is inevitably biased towards depictions in European maritime mapping. A forthcoming section on cetacean stranding introduces the rich heritage of cetaceans in indigenous oral traditions and cosmologies that also expand into larger contexts of human-cetacean relations that complicate the settler/indigenous binaries.

References:
Andrade, Claudia D., Taryn Sepúlveda, Cristóbal Rivera, Cristian Aldea, and Tomás I. Marina. 2026. ‘Marine Trophic Architecture and Hidden Ecological Connections in the Strait of Magellan: Keystone Species and Ecosystem Resilience’. Oikos 2026 (4): e11711. https://doi.org/10.1002/oik.11711.
Arnon, Inbal, Simon Kirby, Jenny A. Allen, Claire Garrigue, Emma L. Carroll, and Ellen C. Garland. 2025. ‘Whale Song Shows Language-like Statistical Structure’. Science 387 (6734): 649–53. https://doi.org/10.1126/science.adq7055.
Baisas, Laura. 2025. ‘Krill or Anchovy? Baleen Whale Songs May Indicate What’s on the Menu’. Popular Science, February 26. https://www.popsci.com/environment/whale-song-food/.
Bakker, Karen. 2024. The Sounds of Life: How Digital Technology Is Bringing Us Closer to the Worlds of Animals and Plants. First paperback printing. Princeton University Press.
Burnett, D. Graham. 2012. The Sounding of the Whale: Science and Cetaceans in the Twentieth Century. University of Chicago Press.
Español‐Jiménez, Sonia, and Mike Van Der Schaar. 2018. ‘First Record of Humpback Whale Songs in Southern Chile: Analysis of Seasonal and Diel Variation’. Marine Mammal Science 34 (3): 718–33. https://doi.org/10.1111/mms.12477.
Ganchrow, Raviv. 2024. ‘Sound’s Spatial-Material Circuitry’. In The Routledge Companion to the Sound of Space, edited by Jane Burry and Emma Kate-Matthews. Routledge.
Gibbons, Jorge, Juan J. Capella, and Carlos Valladares. 2003. ‘Rediscovery of a Humpback Whale (Megaptera Novaeangliae) Feeding Ground in the Straits of Magellan, Chile’. J. Cetacean Res. Manage. 5 (2): 203–8. https://doi.org/10.47536/jcrm.v5i2.818.
Guzman, H. M. 2016. New Routeing Measure Other Than Traffic Separation Schemes, Rules for Navigating Through the Strait of Magellan off Chile. NCSR 4/3/X. Routing Measures And Mandatory Ship Reporting Systems. International Maritime Organization. https://www.directemar.cl/directemar/site/docs/20170301/20170301151332/827_19.pdf.
Hayward, Alexander, Simon W. Wright, Dustin Carroll, et al. 2025. ‘Antarctic Phytoplankton Communities Restructure under Shifting Sea-Ice Regimes’. Nature Climate Change 15 (8): 889–96. https://doi.org/10.1038/s41558-025-02379-x.
Hegeman, Milou, Nicholas B. W. Macfarlane, Philippe Verborgh, et al. 2026. ‘Vocal Compensation to Noise in Long-Finned Pilot Whales ( Globicephala Melas )’. Journal of Experimental Biology 229 (9): jeb251217. https://doi.org/10.1242/jeb.251217.
Mercado, Eduardo. 2018. ‘The Sonar Model for Humpback Whale Song Revised’. Frontiers in Psychology 9 (July). https://doi.org/10.3389/fpsyg.2018.01156.
Nahas Miranda, Michel. 2026. ‘Caída de Más Del 85%: Expertos Alertan Sobre Desaparición Del Pingüino de Magallanes En La Isla Magdalena de Chile’. CNN Chile, January 21. https://www.cnnchile.com/pais/detalles-drastica-caida-colonia-pinguinos-isla-magdalena_20260121/.
Niemeyer, Hans. 2010. Crónica de un descubrimiento. Las pinturas rupestres de El Médano Taltal. Museo Chileno de Arte Precolombino.
Payne, Roger S., and Scott McVay. 1971. ‘Songs of Humpback Whales’. Articles. Science 173 (3997): 585–97. https://doi.org/10.1126/science.173.3997.585.
Ryan, John P., William K. Oestreich, Kelly J. Benoit-Bird, et al. 2025. ‘Audible Changes in Marine Trophic Ecology: Baleen Whale Song Tracks Foraging Conditions in the Eastern North Pacific’. PLOS ONE 20 (2): e0318624. https://doi.org/10.1371/journal.pone.0318624.
Schall, Elena, Karolin Thomisch, Olaf Boebel, Gabriele Gerlach, Sari Mangia Woods, Irene T. Roca, et al. 2021. ‘Humpback Whale Song Recordings Suggest Common Feeding Ground Occupation by Multiple Populations’. Scientific Reports 11 (1): 18806. https://doi.org/10.1038/s41598-021-98295-z.
Schall, Elena, Karolin Thomisch, Olaf Boebel, Gabriele Gerlach, Sari Mangia Woods, Ahmed El-Gabbas, et al. 2021. ‘Multi-Year Presence of Humpback Whales in the Atlantic Sector of the Southern Ocean but Not during El Niño’. Communications Biology 4 (1): 790. https://doi.org/10.1038/s42003-021-02332-6.
Schulze, Josephine N., Judith Denkinger, Javier Oña, M. Michael Poole, and Ellen C. Garland. 2022. ‘Humpback Whale Song Revolutions Continue to Spread from the Central into the Eastern South Pacific’. Royal Society Open Science 9 (8): 220158. https://doi.org/10.1098/rsos.220158.
Shabangu, Fannie W., and Katie A. Kowarski. 2022. ‘The Beat Goes On: Humpback Whale Song Seasonality in Antarctic and South African Waters’. Frontiers in Marine Science 9 (April): 827324. https://doi.org/10.3389/fmars.2022.827324.
Steingo, Gavin. 2024. Interspecies Communication: Sound and Music beyond Humanity. The University of Chicago Press.
Stimpert, Alison K., Lindsey E. Peavey, Ari S. Friedlaender, and Douglas P. Nowacek. 2012. ‘Humpback Whale Song and Foraging Behavior on an Antarctic Feeding Ground’. PLoS ONE 7 (12): e51214. https://doi.org/10.1371/journal.pone.0051214.
Stimpert, Alison K., David N. Wiley, Whitlow W. L. Au, Mark P. Johnson, and Roland Arsenault. 2007. ‘“Megapclicks”: Acoustic Click Trains and Buzzes Produced during Night-Time Foraging of Humpback Whales ( Megaptera Novaeangliae )’. Biology Letters 3 (5): 467–70. https://doi.org/10.1098/rsbl.2007.0281.
Suzuki, Ryuji, John R. Buck, and Peter L. Tyack. 2006. ‘Information Entropy of Humpback Whale Songs’. The Journal of the Acoustical Society of America 119 (3): 1849–66. https://doi.org/10.1121/1.2161827.
Toro, Frederick, Susannah J. Buchan, Mario Alvarado-Rybak, et al. 2025. ‘High Rate of Ship Strike to Large Whales off Chile: Historical Data and Proposed Actions to Reduce Risk’. Marine Policy 174 (April): 106577. https://doi.org/10.1016/j.marpol.2024.106577.
Research support in collaboration with the Institute of Sonology, University of the Arts The Hague and Chair of Transcultural Musicology, Institute of Musicology, Humboldt University Berlin.

Technical support by aporee.org / radio.earth, Triton Audio and Sommer Cable.

Logistics and archipelago access in collaboration with Whale Sound, Punta Arenas / Carlos III.

We would like to personally thank Gökay Atabek, Luis Barrientos, Cameron Bodine, Lex van den Broek, Miguel Caceres, Robert Carracedo Recasens, Florencia Curci, Libby Eyre, Ivan Flores, Michaela Frühwirth, Sila Ganchrow, Alberto Garcia Piquer, Jorge Gibbons, Luis Gonzales, Eduardo Hernandez, Sebastian Klotz, Krista Michelle McGrath, Udo Noll, Jonathan Oberreuter, Christophe Pollet, Alfredo Prieto, Lilian Riquelme, Andrés Ruiz, Simone Sacchi, Guillermina Saldivia Yáñez, Alberto Serrano, Juan Carlos Campos Solari, Nicolas Spencer, Thea Stabenow, Federico Stäger, Carlos Valladares, Simon Valladares, Martin van der Velde.